神奈川県が2003年に作成した《水源涵養域での広葉樹林整備マニュアル》の概要を紹介しながらそこから見えてくる課題を検証してみるシリーズの5回目の今回は、丹沢・大山に拡がる植生=植物群落の実際を見てみることから始めます。
その前にここでもう一度、神奈川県が考えている広葉樹林の整備の基本方針を確認しておきます。主に次の3点にまとめることができます。
● 対象エリア全体を《土壌保全型》と《生産配慮型》の2タイプに分ける。
● 次に(植生の違いに対応して)標高800mを境に全く異なる整備内容にする。
● それぞれ《土壌保全》・《森林管理》・《基礎改良》の3つの整備区分を設ける。
この基本方針に基づいて、整備内容を具体的に落とし込んだものが下の一覧表になります。
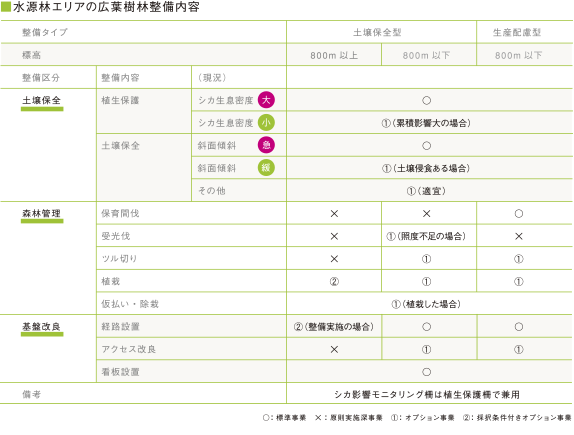 ■
■
この整備内容の一覧から見えてくるのは、標高800m以下での木材生産を希望する私有林である《生産配慮型》区域は、ほぼ人工林整備に準じた内容になっています。またシカの食害からの植生保護および土壌の侵食防止を全エリアで実施することにし、これ以外の《土壌保全型》区域についての《森林管理》は以下の二つにまとめることができます。
● 800m以上では《森林管理》・《基礎改良》に関わる整備は行わず、自然に任せる。
● 800m以下では《基礎改良》整備を行うが、《森林管理》は必要な場合に限る。
以上のことから、元来800m以下の潜在自然植生である常緑広葉樹林に替わり、階層構造が豊かな落葉広葉樹林を人工的に育て上げるためには、どうしても整備が必要になることを示しています。自然に任せるとヒトの手をかけずに成長するであろう、この区域の潜在自然植生である常緑広葉樹林では何故ダメなのか?その理由はシリーズの第2回で書いたように「潜在自然植生である常緑広葉樹林は下層植生が貧弱」なために、水源林としては適正を欠くという県の主張でした。
そこで、この私にとっては納得できない指摘に対して神宮の森なども一例に挙げながら、そうでもない場合があることを宮脇さんの著作を引用することで、私なりのオブジェクションを試みてきたのですが、今回は、少し趣向を変えて、丹沢・大山に拡がる植生=植物群落の実際を見てみようというわけです。
さっそく標高800m以下の地域の植生を網羅的に調査した神奈川県の『丹沢大山総合調査学術報告書』(2007年)から該当部分を写し取ってみましょう。まずは、この地域にわずかに残存するとされているヤブツバキクラスの常緑広葉樹林の現状です。
<常緑広葉樹林: ヤブツバキクラス>
丹沢大山の常緑広葉樹林(ヤブツバキクラス) は、今回の調査では、ヤブコウジ-スダジイ群集、サカキ-ウラジロガシ群集、イロハモミジ-ケヤキ群集、アラカシ群落の3群集1群落にまとめられた。これらの常緑広葉樹林の多くは標高700~800m以下の山地に分布している。西丹沢、大山などの一部地域では標高900~1,000mの尾根や南向き斜面にヤブツバキクラスの常緑植物を伴うアカガシ林が分布しており、局所的なヤブツバキクラス域の上昇が認められる。現在、丹沢大山のヤブツバキクラス域はスギ・ヒノキ植林が広く占めており、常緑広葉樹林はごく一部の尾根などに残存するにすぎない。面積的にまとまりのある常緑広葉樹林は大山、札掛、大洞沢、境沢、本谷川周辺のモミ-カシ林(サカキ-ウラジロガシ群集)が挙げられる。これらの地域では近年、 モミの衰弱、枯死が進行しており、今後の動態に注視する必要がある。また、ブナクラスの森林群落と同様に、丹沢大山の常緑広葉樹林においても総じてシカ(一部イノシシ)による林床植生の強度のかく乱(喫食・踏み付け)が認められる。 低木層、草本層の退行に伴う土壌流出や森林の更新阻害など多くの問題が懸念される。丹沢大山地域の常緑広葉樹林は、標高700~800m以下の低標高域において野生生物の環境収容力を適正に保持するための鍵となる植生でもあり、その保全・再生に積極的に努めてゆく必要がある。
このように、3群集+1群落の常緑広葉樹林がスギ・ヒノキ植林の間隙を縫うように、ごく一部の尾根などに残存しているにすぎません。しかもシカ(一部イノシシ)によるかく乱が深刻であると報告されており、もはや丹沢大山の常緑広葉樹林は風前の灯びという状況のように思えます。おそらく、神奈川県の「常緑広葉樹林は下層植生が貧弱」という指摘はこの状況に対する直接的な反応だと考えられなくもなく、その限りでは仕方がないことなのでしょうか。
続いて、3群集+1群落の詳細をそれぞれ見てみましよう。
● ヤブコウジ-スダジイ群集
1952標徴種・区分種: スダジイ、ヤブニッケイ、ヒサカキ、ツルグミなど
日向山山頂付近の南向き尾根斜面(伊勢原市、海抜380m)には、高木層にウラジロガシ、アカガシ、モミの混生したスダジイ優占林が生育している。このスダジイ林はヤブコウジ-スダジイ群集にまとめられた。調査林分は植生高18~20m、高木層から草本層までの4層からなり、スダジイ、シロダモなどのヤブツバキクラスの常緑植物を主体に15~17 種が出現する。林内の低木層、草本層はそれぞれ10~20%、1%のまばらな植被からなり、シキミ、ヒイラギ、テイカカズラ、キヅタ、ホウライカズラ、タブノキなどのヤブツバキクラスの常緑植物が優占度 +~1で生育する。日向山の林分はシカによる喫食、踏み付けなどの影響で低木層・草本層が貧化した退行相と判定される。林内では表層土壌の浸食・流出が懸念されるため、今後の動態を注視し、植生・立地の保全を検討する必要がある。丹沢山地とその県内周辺では、本報の日向山のほか、大井町(海抜100m付近)に本群集の記録(神奈川県教育委員会、1972)がある。日向山の調査林分は、県内における本群集域の上限(より高海抜側に分布するサカキ-ウラジロガシ群集域との境界域) に位置する数少ないスダジイ林として学術的に重要である。● サカキ-ウラジロガシ群集
1949標徴種・区分種: スズダケ、アセビ、クロモジ、ツクバネウツギなど
ウラジロガシ、アカガシ、ツクバネガシ、モミを主体とする常緑広葉樹林。調査資料は海抜435~985mから得られている。調査林分は植生高13~ 35m、高木層から草本層までの4層からなり、ウラジロガシ、アカガシ、モミ、シキミ、ヒイラギなどのヤブツバキクラスの常緑植物を主体に計20~39種が出現する。林内の低木層、草本層はそれぞれ10~30%、5~30%のまばらな植被で、シキミ、カヤ、ウラジロガシ、テイカカズラ、スズダケ、アセビ、ヒメカンスゲなどの常緑植物などが優占度 +~ 3で生育する。サカキ-ウラジロガシ群集は今回の調査では、下位単位として、コカンスゲ、ノササゲ、シュンランなどを区分種とするシュンラン亜群集とモミ、イヌシデ、キッコウハグマ、キブシを区分種とするモミ亜群集が区分された。シュンラン亜群集はアカガシ、ツクバネガシの優占林で、崩壊を伴わない安定した山地中庸~尾根斜面などに生育する。モミ亜群集はモミの優占林で、シカによる過度の喫食圧がかかる東丹沢の林分がまとめられた。このモミ亜群集では、シカによる喫食、踏み付けの痕跡、シカの嗜好性の低いシキミやアセビの増加、嗜好性の高いスズダケの退行が確認される。
■
現在、丹沢山地で見られるサカキ-ウラジロガシ群集は、札掛や大山南面などの比較的まとまりのある林分を除き、急傾斜地の残存林であることが多い。シカの影響により下層植生の退行した林分では表層土壌の浸食・流出が懸念されるため、今後の動態に注視する必要がある。
■
本群集は、海抜700~ 800m以下の山地中庸~尾根斜面を生育域とする、丹沢山地ヤブツバキクラスの中核をなす植生タイプであるが、現在その生育域はほとんどがスギ・ヒノキ植林に置き換えられている。残存林としては、布川、本谷川、大洞沢、境沢流域、札掛の学術考証林(清川村)および大山南斜面(伊勢原市)に発達した林分が認められる他は、断片的なものが多い。今回大山南斜面で調査された海抜900~1,000mの南向き尾根斜面に成立したアカガシ優占林(一部モミが混生)は県内ヤブツバキクラス域の上限に位置する極相林の一タイプと判定される。このようなアカガシ優占林はしばしばブナクラスの夏緑広葉樹を多く伴うことから、組成的にコカンスゲ-ツガ群集などのブナクラス植生の下位単位として扱われることもある。本報告ではサカキ-ウラジロガシ群集に含められた。● イロハモミジ-ケヤキ群集
1970標徴種・区分種: ケヤキ, イロハモミジ, コゴメウツギ, サワシバ
中津川、布川沿いの渓谷斜面には、ウラジロガシ、カヤなどのヤブツバキクラスの常緑広葉樹を主体に、ケヤキ、イロハモミジ、エゾエノキ、クマノミズキ、ミズキ、ミヤマハハソなどの夏緑広葉樹が混生した渓谷林が生育している。この渓谷林はイロハモミジ-ケヤキ群集にまとめられた。 調査資料は海抜435~500mの林分から得られた。調査林分は植生高15~17m、高木層から草本層までの4層からなり、林内の低木層、草本層はそれぞれ植被率10~20%、1~10%と疎らで、シキミ、アセビ、カヤなどの常緑植物やスズダケ、コゴメウツギ、ヤマブキなどが優占度+~1で生育する。出現種数は24~39種である。調査林分の生育立地は、排水良好な適潤土壌の堆積した渓谷斜面である。より岩礫に富み、崩壊・崩積が繰り返される不安定な立地では、タマアジサイ-フサザクラ群集が発達する。調査林分ではシカによる喫食・踏み付けの痕跡、シカの嗜好性の低いシキミやアセビの増加、嗜好性の高いスズダケの退行が確認される。現在、丹沢山地で見られるイロハモミジ-ケヤキ群集は河川、沢沿いの傾斜地の残存林であることが多く、シカの影響で下層植生が退行した場合、表層土壌が容易に浸食・流出する点が懸念される。イロハモミジ-ケヤキ群集は、 丹沢山地のヤブツバキクラス域における代表的なケヤキ林であるが、その植生域は主としてスギ・ヒノキ植林に置き換えられている。● アラカシ群落
区分種: アラカシ, アズマネザサ
県道710号線が貫く中津川沿いの渓谷斜面( 海抜240m)には、岩礫混じりの浅土地にアラカシ優占林が発達している。このアラカシ林は、人里近くに生育する二次的なアラカシ優占林(伊勢原市高橋沢、海抜190m)とともに、アラカシ群落にまとめられた。調査林分は、植生高13~17m、高木層から草本層までの4層構造を持つ。高木層、亜高木層にはアラカシが優占し、渓谷斜面生の林分ではケヤキ、エノキが混生する。低木層、草本層にはアオキ、キヅタ、シロダモなどのヤブツバキクラスの常緑植物などが出現する。丹沢山地のヤブツバキクラス域下部の渓谷沿いの岩角地が本群落の潜在的な生育域と推定される。本群落のようなアラカシ優占林は、丹沢山地では酒匂川、皆瀬川、早戸川などの渓谷沿いでよく観察される。このうち、早戸川沿いに見られる傾斜30°以上の空中湿度の高い渓谷斜面、岩角地のアラカシ林は、オオバジャノヒゲ-アラカシ群集とみなされている(国際生態学センター、未発表)。今回の調査では資料数が少なく、他のヤブツバキクラスの群集と本群落との組成的な差が明瞭でないため、群集帰属を保留した。
ヤブコウジ-スダジイ群集およびサカキ-ウラジロガシ群集はともに丹沢のヤブツバキクラスを代表する植生ですが、このようにその大部分はスギ・ヒノキ植林に置き換えられ、加えてシカの食害なども重なり、退行相の様相を呈しているそうです。
注目すべきはイロハモミジ-ケヤキ群集です。ウラジロガシ、カヤという常緑広葉樹を主体にケヤキ、イロハモミジなどの夏緑広葉樹が混生した植生が出てきました。丹沢のヤブツバキクラス域における代表的なケヤキ林であるとされています。実際には(本などの情報源では知ることができないような)常緑と夏緑が混生した広葉樹があることがわかってきました。これも他の二つの群集と同じように、その植生域は今ではスギ・ヒノキ植林に置き換えられているとあります。
次に、標高800m以下の地域で県が整備の対象とする夏緑(落葉)広葉樹林を見てみましょう。まずは全標高域の概要から。
<夏緑広葉樹林: ブナクラスほか>
丹沢山地における森林帯の垂直分布は、渓谷沿いや尾根部など地形によって多少変動するものの、 海抜 600 ~700m を境界として下部の常緑広葉樹林域: ヤブツバキクラス域から夏緑広葉樹林域: ブナクラス域に移行する。 ブナクラス域の森林植生は広葉樹林が広く残されているが、スギ、 ヒノキの人工林も同様に広い面積を占めている。また清川村札掛、大洞などにはモミ、ツガなどの常緑針葉樹林が尾根部を中心に残存している。 夏緑広葉樹林の自然林ではブナ林が海抜 1,000m 以上の地域に広く残されているが、 喫食、 踏つけなどシカによる林床植生のかく乱が進み、深刻な事態となっている。海抜 1,100m 以下の斜面下部や谷沿いの急斜面には局所的にイヌブナ林がみられ、その周辺にはクマシデ、イヌシデなどのシデ林が二次林あるいは持続群落として生育している。二次林ではかつて薪炭林として利用されていたコナラ林やミズナラ林がみられるが、面積的には少ない。沢沿いの湿生林は、ブナクラス域下部にはケヤキ林が生育し、ほぼ海抜 800m 以上にはサワグルミ林、シオジ林が分布している。 また、各地の沢沿い崩壊地にはフサザクラを主体とする先駆群落がみられる。
この調査報告書では、丹沢でのヤブツバキクラス域とブナクラス域の境界を600〜700mとしていますが、ヤブツバキクラス域では薪炭林としてのコナラ林やミズナラ林が二次林あるいは持続群落として残っているとあります。これらヤブツバキクラス域ニアる夏緑(落葉)広葉樹林の群集・群落を詳しく見てみます。
●クリ-コナラ群集
標徴種・区分種: コナラ, ヤマツツジ, エゴノキ, コゴメウツギ, コウヤボウキなど
コナラを主体とする夏緑広葉樹林はクリ-コナラ群集にまとめられた。 クリ-コナラ群集は、 薪炭林、 農用林として利用管理されてきた二次林で、 海抜280mのヤブツバキクラス域から海抜 920mのブナクラス域下部にかけての範囲から植生調査資料が得られた。 群落高は10~15m前後で、3~ 4層構造の植分が多い。 高木層にはコナラが優占することが多いが、 ブナクラス域では時にミズナラが優占する。 優占種のほかエンコウカエデ、 クマシデ、イヌシデ、ヤマザクラ、 エゴノキ、アオハダなど多くの夏緑広葉樹が混生している。低木層には、ムラサキシキブ、 クロモジ、ダンコウ バイ、コゴメウツギ、ウリカエデ、ガマズミ、 ヤマツツジ、ヤマボウシなどのほか、ヤブツバキクラス域では常緑広葉樹のシロダモ、 アオキもみられる。草本層はコウヤボウキ、コ チヂミザサ、フジ、タチツボスミレ、ヤブレガサ、ナガバジャノヒゲなどのほか多くの林床植物が生育している。 中には草本層の植被率は 5 ~ 10% に過ぎなくても 40 種を超える林床植物を持つ植分もみられる。このような植分では林床植物のサイズが矮小化しているのが特徴で、 シカの喫食圧と蹄による表層土のかく乱が強く影響していると考えられる。また、ブナクラス域ではスズダケが密生することがあり、そのような植分では15種前後に過ぎない。調査されたコナラ林は、全国のコナラ林をまとめた総合常在度表の原表と比較した結果、クリ-コ ナラ群集に同定された。しかし、本報のクリ-コナラ群集は他地域のクリ-コナラ群集の標徴種あるいは常在度が高いとされているオトコヨウゾメ、 ハクウンボク、 チョウジザクラ、アワブキ、ヤマガシュウなど種を欠くか常在度が低い。これ にはシカによるかく乱が関係していると推定される。 また、 今回まとめられたクリ-コナラ群集はミズナラとスズダケが優占あるいは混生する植分をもち、太平洋側のコナラ林としては相観的にミズナラ林に類似した特徴をもっている。
■
●アカシデ-クマシデ群落
区分種: アカシデ、ヒメノガリヤス、ヤマカモジグサ、トウゴクミツバツツジ、ホソエカエデ、タマアジサイなど 山腹の谷に面した斜面上部から尾根肩部にかけての急傾斜地あるいは沢沿いの崩壊性の強い斜面には、 イヌブナ林あるいはミズナラ林に隣接してクマシデ、アカシデ、イヌシデ、サワシバなどのシデ類を主体とする夏緑広葉樹林が生育している。これらのシデ林はアカシデ-クマシデ群落 にまとめられた。アカシデ-クマシデ群落は、 高さ 15m前後の4層群落が多いが10m以下の3層群落もみられる。 高木層はクマシデが優占する傾向があるが、 植被の大半を占めることは稀で、多くの場合シデ類のイヌシデ、サワシバのほか、 ヨグソミネバリ、オオモミジ、 ヤマハンノキ、ミヤマヤシャブシなどが混生して林冠を構成している。クリ-コナラ群集とフクオウソウ-ミズナラ群集に対しては、 相観的には高木層 にミズナラを伴わないことにより識別可能である。 亜高木層は高木層との共通種で構成される。 低木層の構成種は、5~10種、平均7種であるが、 2つ以上の植分に共通してみられる種はウツギ、トウゴクミツバツツジ、アセビ、スズダケのみで、植分間の共通種は少ない。それに対し草本層にはきわめて多くの種が生育しており、ヒメノガリヤス、ヤマカモジグサ、キッコウハグマ、モミジイチゴ、スズダケ、マメザクラ、 シロヨメナなど平均29種を数える。また、林床植物はシカの食害を受けている個体が多くみられる。アカシデ-クマシデ群落は、急斜面に生育していたイヌブナ林やブナ林の二次林と考えられる。土壌中に角礫の多い。 あるいは露頭がみられる表層土が移動しやすい不安定な立地に生育しており、 丹沢山地における斜面保全上 の重要な役割を果たしている。
■
●フクオウソウ-ミズナラ群集
標徴種: イトマキイタヤ
区分種: ヤマムグラ、 シロイトスゲ、 ハナタデ、タニタデ、ヘビノネゴザ、コハウチワカエデなど
フクオウソウ-ミズナラ群集はヤマムグラ、シロイトスゲなどの多くの種により区分される夏緑広葉樹二次林である。植生高15~22mの高木林であるが、亜高木層を欠くことも多い。 高木層はミズナラ、 イヌシデ、ミズキ、 イトマキイタヤなど8~9種もの樹種が生育するが、優占種はミズナラ、 エンコウカエデなど植分により異なっている。 低木層にはアブラチャン、 ツクバネウツギ、 カマツカ、 ガマズミ、ヤマグワなどがみられる。 草本層には上述の区分種のほか、イワガラミ、 ツルマサキ、 ムカゴイラクサ、テンニンソウ、コチヂミザサ、アシボソ、ヘビノネゴザなど平均39種ものきわめ て多くの種が生育している。また、草本層はミズナラ、 サンショウ、ムラサキシキブ、アオダモ、ウツギ、キブシ、タラノキ、 モミジイチゴなど木本類の常在度が高いのが特徴である。フクオウソウ-ミズナラ群集はアカシデ-クマシデ群落と同様に、ブナ林やイヌブナ林の代償植生と考えられるが、よ り表層土壌の厚い安定した立地に生育している。 海抜980~1,290mの地域から得られた 3 植分がまとめられている。 群落区分種のハナタデ、タニタデ、アシボソ、 マツカゼソウ、クワガタソウ、ヒメチドメ、ヤマミズなどはシカによる食害や撹乱を強く受けている森林の林床に特徴的な種で、シカの影響により生じた種群と考えられる。また、 出現一回のみ の種も多く、種組成的なまとまりを欠いている。丹沢山地を含めた神奈川県のミズナラ林の群落分類は、これまでまとまった研究報告に乏しく、 その実態は明らかではなかった。全国のミズナラ林群落をまとめた においても神奈川県のミズナラ林についてはほとん ど触れられていない。しかし、 今回のデータを全国のミズナラ林の総合常在度表に組み込み比較したところ、 シカの食害による影響に起因すると考えられる種組成的貧化や多くのシカの非嗜好性植物の存在など種組成的 まとまりに欠ける傾向がみられるものの、フクオウソウ-ミズ ナラ群集の標徴種であるイトマキイタヤ、マメザクラを有することから、 中部地方太平洋側を中心に分布域を持つ同群 集に所属された。
■
●コカンスゲ-ツガ群集
標徴種・区分種: モミ、 イヌブナ、コカンスゲ、ツガ、 アワブキ、 コバノガマズミ、ウラジロガシ、テイカカズライヌブナはブナクラス域下部でしばしばモミやツガと混生し、 急斜面、 尾根などの表層土の浅い不安定立地における土地的な自然林を形成する。丹沢山地でも急峻な不安定斜面や尾根を中心にイヌブナやモミあるいはツガの優占または混生する植分がみられる。これらの植分は上の標徴種・区分種によりコカンスゲ-ツガ群集にまとめられた。
コカンスゲ-ツガ群集は、高木層にイヌブナ、 クマシデ、アカシデ、ブナなどの夏緑広葉樹やウラジロガシ、アカガシなどの常緑広葉樹のほか常緑針葉樹のツガ、モミなど多くの樹種が生育している。これらの樹種は植分により優占度が異なることが多く、イヌブナ林、クマシデ林、ブナ林、コナラ林、モミ-ツガ林など多様な群落相観を形成している。
群落構造は高さ10mあまりの3層群落もみられるが、 多くの植分では20mを超える4層群落で、発達した植分では胸高直径1m前後で高さ25m以上に達するモミ-ツガ林もみられる。 高木層には上述の種のほかにヨグソミネバリ、 ミズナラ、 エンコウカエデ、 ヤマザクラ、カヤなども混生している。 亜高木層にはイヌブナ、サワシバ、オオモミジ、ウラジロガシ、アカガシ、カヤなどが混生する。 低木層には常在度の高い種は少ないが、モミ、イヌブナ、カヤなど高木層構成種の幼樹をはじめ、ムラサキシキブ、コバノガマズミ、ミツバツツジ、ツクバネウツギ、ヤブムラサキ、 アブラチャンなど多くの夏緑低木類から構成される。草本層はスズダケの常在度がコカンスゲと共に高く、優占している植分も多い。
植被率は植分による差が大きく、スズダケが優占している植分では40~90%と高いが、シカによる食害を強く受けている植分では低い傾向がみられ、特に低海抜のシキミ亜群集では3~5%ときわめて低くなっている。
コカンスゲ-ツガ群集は、垂直分布的にはヤマボウシ-ブナ群集の下方に分布域をもつコナラ-ミズナラオーダーの植生として位置づけられる。 標高や地域の違いにより以下の3下位単位に区分される。 シキミ亜群集は、ツガ、 シキミ、イロハモミジ、サルトリイバラ、マルバウツギにより区分される、 海抜490~780mのヤブツバキクラス域を含む低海抜地生の植分である。ウラジロガシ、シキミ、アラカシなどヤブツバキクラスの種を含み、イヌブナは優占せず、モミ、ツガ、 ウラジロガシが優占するなど種組成的、相観的に常緑広葉樹林のサカキ-ウラジロガシ群集に類似している。しかし、シキミ亜群集は多くのブナクラスの標徴種を伴うことで区分される。特別の区分種を持たない典型亜群集は、イヌブナ、クマシデが優占するシデ-イヌブナ林で、西丹沢の植分がまとめられている。ブナ亜群集は、ブナ、ミツバツツジ、イワガラミ、タンナサワフタギ、バイカツツジ、カンスゲにより区分される。コカンスゲ-ツガ群集の中でも海抜810~1,070mの高海抜地に生育している。ブナ亜群集はブナの優占度が高く、 林床にスズダケが優占するなど北関東から東北地方太平洋側に分布するとされているブナ-イヌブナ群集に類似している。丹沢山地において、ブナ亜群集に対応すると考えられるイヌブナを含むブナ林をブナ-イヌブナ群集にまとめ、 ヤマボウシ-ブナ群集の下部に分布するブナ林としてコナラ-ミズナラオーダーに位置づけている。 コカンスゲ-ツガ群集とブナ-イヌブナ群集については、種組成的、相観的および生態的共通性が高いが、これまで両者の種組成比較が詳細に行われていないため混乱がある。本報告では、ブナ亜群集はブナを含まないシキミ亜群集や典型亜群集とともに、上述の標徴種・区分種をササ-ブナオーダーのヤマボウシ-ブナ群集, オオモミジガサ-ブナ群集との明確な対立種群としてもつことから、イヌブナ林の群集として扱った。
以上、3群集1群落の夏緑(落葉)広葉樹林紹介しましたが、280mのヤブツバキクラス域〜920mのブナクラス域にまたがって分布するクリ—コナラ群集以外はいづれも広域に生育するものや急峻で不安定な斜面や谷間、尾根を中心に分布しているようです。これらは報告書の記述にもあるように、イヌブナやブナ林の二次林であったり、その代償植生であるとみられています。
(続く)
![]()

